WikiJournal of Science/Arabinogalactan-proteins
![]()
WikiJournal of Science
Open access • Publication charge free • Public peer review • Wikipedia-integrated
This article has been through public peer review.
Post-publication review comments or direct edits can be left at the version as it appears on Wikipedia.First submitted:
Accepted:
Reviewer comments
PDF: Download
DOI: 10.15347/WJS/2021.002
QID: Q99557488
XML: Download
Share article
![]() Email
|
Email
| ![]() Facebook
|
Facebook
| ![]() Twitter
|
Twitter
| ![]() LinkedIn
|
LinkedIn
| ![]() Mendeley
|
Mendeley
| ![]() ResearchGate
ResearchGate
Suggested citation format:
Yingxuan Ma; Kim Johnson (15 January 2021). "Arabinogalactan-proteins". WikiJournal of Science 4 (1): 2. doi:10.15347/WJS/2021.002. Wikidata Q99557488. ISSN 2470-6345. https://upload.wikimedia.org/wikiversity/en/0/09/Arabinogalactan-proteins.pdf.
Citation metrics
AltMetrics
Page views on Wikipedia
Wikipedia: Content from this work is used in the following Wikipedia article: Arabinogalactan protein.
License: ![]()
![]() This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction, provided the original author and source are credited.
This is an open access article distributed under the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction, provided the original author and source are credited.
Dan Graur ![]() (handling editor) contact
(handling editor) contact
Florian Weller ![]() contact
contact
Milan Dragićević ![]()
Elisabeth Jamet
Azeddine Driouich ![]()
Article information
Authors: Yingxuan Ma[a], Kim Johnson[a][i] ![]()
- ↑ 1.0 1.1 School of BioSciences, University of Melbourne, Parkville, Victoria 3010, Australia;
La Trobe Institute for Agriculture & Food, Department of Animal, Plant and Soil Sciences, La Trobe University, Bundoora, Victoria 3086, Australia;
Sino-Australia Plant Cell Wall Research Centre, State Key Laboratory of Subtropical Silviculture School of Forestry and Biotechnology, Zhejiang A&F University, Hangzhou, 311300, China
- ↑ k.johnson@latrobe.edu.au
Abstract
AGPs have been reported in a wide range of higher plants in seeds, roots, stems, leaves and inflorescences. They have also been reported in secretions of cell culture medium of root, leaf, endosperm and embryo tissues, and some exudate producing cell types such as stylar canal cells are capable of producing lavish amounts of AGPs.
AGP protein backbones and classification

The protein component of AGPs is rich in the amino acids Proline (P), Alanine (A), Serine (S) and Threonine (T), also known as ‘PAST’, and this amino acid bias is one of the features used to identify them.[1][2][3][4][5] AGPs are intrinsically disordered proteins as they contain a high proportion of disordering amino acids such as Proline that disrupt the formation of stable folded structures. Characteristic of intrinsically disordered proteins, AGPs also contain repeat motifs and post-translational modifications.[2][6] Proline residues in the protein backbone can be hydroxylated to Hydroxyproline (O) depending on the surrounding amino acids. The ‘Hyp contiguity hypothesis’ [7][2][3] predicts that when O occurs in a non-contiguous manner, for example the sequence 'SOTO', such as occurs in AGPs, this acts as a signal for O-linked glycosylation of large branched type II arabinogalactan (AG) polysaccharides.[8] Sequences that direct AG glycosylation (SO, TO, AO, VO) are called AGP glycomotifs (Figure 1).
All AGP protein backbones contain a minimum of 3 clustered AGP glycomotifs and an N-terminal signal peptide that directs the protein into the endoplasmic reticulum (ER) where post-translational modifications begin.[9] Prolyl hydroxylation of P to O is fulfilled by prolyl 4-hydroxylases (P4Hs) belonging to the 2-oxoglutarate dependant dioxygenase family.[10] P4H has been identified in both the ER and Golgi apparatus (GA).[11] The addition of the glycosylphosphatidylinositol (GPI)-anchor occurs in most but not all AGPs.[3][4]
AGP family of glycoproteins
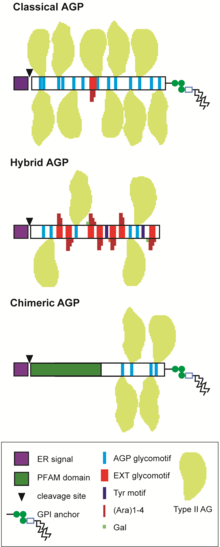
AGPs belong to large multigene families and are divided into several sub-groups depending on the predicted protein sequence.[12][4][13][14][2][3][15] "Classical" AGPs include the GPI-AGPs that consist of a signal peptide at the N-terminus, a PAST-rich sequence of 100-150 aa and a hydrophobic region at the C-terminus that directs addition of a GPI-anchor; non GPI-AGPs that lack the C-terminal GPI signal sequence, Lysine(K)-rich AGPs that contain a K-rich region within the PAST-rich backbone and AG-peptide that have a short PAST-rich backbone of 10-15 aa (Figure 2). Chimeric AGPs consist of proteins that have an AGP region and an additional region with a recognised protein family (Pfam) domain. Chimeric AGPs include fasciclin-like AGPs (FLAs), phytocyanin-like AGPs (PAGs/PLAs, also known as early-nodulin-like proteins, ENODLs) and xylogen-like AGPs (XYLPs) that contain lipid-transfer-like domains.[1] Several other putative chimeric AGP classes have been identified that include AG glycomotifs associated with protein kinase, leucine-rich repeat, X8, FH2 and other protein family domains.[15][16][17] Other non-classical AGPs exist such as those containing a cysteine(C)-rich domain, also called PAC domains, and/or histidine(H)-rich domain,[18][19] as well as many hybrid HRGPs that have motifs characteristic of AGPs and other HRGP members, usually extensin and Tyr motifs.[18][20][1][2][3] AGPs are evolutionarily ancient and have been identified in green algae as well as Chromista and Glaucophyta.[2][3][21] Found throughout the entire plant lineage, land plants are suggested to have inherited and diversified the existing AGP protein backbone genes present in algae to generate an enormous number of AGP glycoforms.
AGP biosynthesis
After translation, the AGP protein backbones are highly decorated with complex carbohydrates, primarily type II AG polysaccharides.[22] The biosynthesis of the mature AGP involves cleavage of the signal peptide at the N-terminus, hydroxylation on the P residues and subsequent glycosylation and in many cases addition of a GPI-anchor.
The structure of the AG glycans consists of a backbone of β-1,3 linked galactose (Gal), with sidechains of β-1,6 linked Gal and have terminal residues of arabinose (Ara), rhamnose (Rha), Gal, fucose (Fuc), and glucuronic acid (GlcA). Glycosylation of the AGP backbone is suggested to initiate in the ER with the addition of first Gal by O-galactosyltransferase, which is predominantly located in ER fractions. [23] Chain extension then occurs primarily in the GA.[24] The AG glycan moiety of AGPs is assembled by glycosyltransferases (GTs).[25] O-glycosylation of AGPs is initiated by the action of Hyp-O-galactosyltransferases (Hyp-O-GalTs) that add the first Gal onto the protein. The complex glycan structures are then elaborated by a suite of glycosyltransferases, the majority of which are bio-chemically uncharacterized. The GT31 family is one of the families involved in AGP glycan backbone biosynthesis.[26][27] Numerous members of the GT31 family have been identified with Hyp-O-GALT activity[28][29] and the core β-(1,3)-galactan backbone is also likely to be synthesized by the GT31 family.[27] Members of the GT14 family are implicated in adding β-(1,6)- and β-(1,3)-galactans to AGPs.[30][31] In Arabidopsis, terminal sugars such as fucose are proposed to be added by AtFUT4 (a fucosyl transferase) and AtFUT6 in the GT37 family [32][33] and the terminal GlcA incorporation can be catalysed by the GT14 family.[30][34] A number of GTs remain to be identified, for example those responsible for terminal Rha.
Bioinformatic analysis predicts the addition of a GPI-anchor on many AGPs.[4] The early synthesis of the GPI moiety occurs on the ER cytoplasmic surface and subsequent assembly take place in the lumen of the ER. These include the assembly of tri-mannose (Man), galactose, non-N-acetylated glucosamine (GlcN) and ethanolamine phosphate to form the mature GPI moiety.[35][36] AGPs undergo GPI-anchor addition while co-translationally migrating into the ER and these two processes finally converge. Subsequently, a transamidase complex simultaneously cleaves the core protein at the C-terminus when it recognizes the ω cleavage site and transfers the fully assembled GPI-anchor onto the amino acid residue at the C-terminus of the protein. These events occur prior to prolyl hydroxylation and glycosylation.[37][11] The core glycan structure of GPI anchors is Man-α-1,2-Man-α-1,6-Man-α-1,4-GlcN-inositol (Man: mannose, GlcN: glucosaminyl), which is conserved in many eukaryotes.[36][38][39][35][11][40] The only plant GPI anchor structure characterized to date is the GPI-anchored AGP from Pyrus communis suspension-cultured cells.[35] This showed a partially modified glycan moiety compared to previously characterized GPI anchors as it contained β-1,4-Gal. The GPI anchor synthesis and protein assembly pathway is proposed to be conserved in mammals and plants.[11] The integration of a GPI-anchor enables the attachment of the protein to the membrane of the ER transiting to the GA leading to secretion to the outer leaflet of the plasma membrane facing the wall.[41] As proposed by Oxley and Bacic,[35] the GPI-anchored AGPs are likely released via cleavage by some phospholipases (PLs) (C or D) and secreted into the extracellular compartment.
AGPs functional roles
Human uses of AGPs include the use of Gum arabic in the food and pharmaceutical industries because of natural properties in thickening and emulsification.[42][43] AGPs in cereal grains have potential applications in biofortification,[44] as sources of dietary fibre to support gut bacteria[45] and protective agents against ethanol toxicity.[46]
AGPs are found in a wide range of plant tissues, in secretions of cell culture medium of root, leaf, endosperm and embryo tissues, and some exudate producing cell types such as stylar canal cells.[20] [47] AGPs have been shown to regulate many aspects of plant growth and development including male-female recognition in reproduction organs, cell division and differentiation in embryo and post-embryo development, seed mucilage cell wall development, root salt tolerance and root-microbe interactions (see Table 1).[5][11][48] These studies suggest that they are multifunctional, similar to what is found in mammalian proteoglycans/glycoproteins.[49][50][51] Conventional methods to study functions of AGPs include the use of β-glycosyl (usually glucosyl) Yariv reagents and monoclonal antibodies (mAbs). β-Glycosyl Yariv reagents are synthetic phenylazo glycoside probes that specifically, but not covalently, bind to AGPs and can be used to precipitate AGPs from solution.[52] They are also used commonly as histochemical stains to probe the locations and distribution of AGPs.[53][54] A number of studies have shown that addition of β-Yariv reagents to plant growth medium can inhibit seedling growth, cell elongation, block somatic embryogenesis and fresh cell wall mass accumulation.[55][56][57] The use of mAbs that specifically bind to carbohydrate epitopes of AGPs have also been employed to infer functions based on the location and pattern of the AGP epitopes.[58] Commonly used mAb against AGPs include CCRC-M7, LM2, JIM8, JIM13 and JIM14.[59]
The function of individual AGPs has largely been inferred through studies of mutants. For example, the Arabidopsis root-specific AtAGP30 was shown to be required for in vitro root regeneration suggesting a function in regenerating the root by modulating phytohormone activity.[60] Studies of agp6 and agp11 mutants in Arabidopsis have demonstrated the importance of these AGPs to prevent uncontrolled generation of the pollen-grain and for normal growth of the pollen-tube.[61][62] The functional mechanisms of AGPs in cell signalling is not well understood. One proposed model suggests AGPs can interact and control the release of calcium from AG glycan (via GlcA residues) to trigger downstream signalling pathways mediated by calcium.[63][64][65] Another possible mechanism, largely based on the study of FLAs, suggests the combination of fascicilin domain and AG glycans can mediate cell-cell adhesion.[66][67] Functions attributed to AGPs are outlined in Table 1.
| Biological role | AGP [a][b] | Location(s) | Function(s) | References |
|---|---|---|---|---|
| Embryogenesis | GhPLA1 | Somatic embryos | Promoting somatic embryogenesis | [68] |
| DcAGPs | Somatic embryos | Promoting somatic embryogenesis | [69] | |
| AtAGPs | Embryos | Embryo development and differentiation | [70] | |
| NtAGPs | Embryos | Embryo development | [71] | |
| BgAGPs | Somatic embryos | Somatic embryo development rate and morphology | [72] | |
| BnAGPs | Embryos | Embryo development | [54] | |
| MaAGPs | Somatic embryos | Promoting somatic embryogenesis | [73] | |
| PsAGPs | Promoting somatic embryogenesis | [74] | ||
| FsAGPs | Embryos | Establishment and stability of the cell wall | [21] | |
| VcALGAL-CAM | embryos | Embryo cell adhesion | [66] | |
| VcISG | embryos | Embryo inversion | [75] | |
| Reproduction | AtAGP4 (JAGGER) | Pistil | Pollen tube blockage | [76] |
| AtAGP6, AtAGP11 | Stamen, pollen grain and pollen tube | Pollen grain development and pollen tube growth | [77][78] | |
| AtAGP18 | Ovule | Megaspore selection | [79][80] | |
| AtFLA3 | Pollen grain and pollen tube | Microspore development | [81] | |
| AtENODL11-15 | Micropylar | Pollen tube reception | [82][83] | |
| BcmMF8 | Pollen grain and pollen tube | Pollen wall development and pollen tube growth | [84] | |
| BcmMF18 | Pollen grain | Pollen grain development, intine formation | [85] | |
| NtTTS | Pistil | Pollen tube growth and guidance | [86] | |
| Np/Na120kD | Pistil | S-specific pollen rejection (self-incompatibility) | [87] | |
| OsMTR1 | Male reproductive cells | Anther development and pollen fertility | [88] | |
| Plant development | AtAGP19 | Stem, flower, root and leaf | Cell division and expansion, leaf development and reproduction | [89] |
| AtAGP57C | Rosette leaf, silique, seed, flower, and shoot apex of inflorescence stem | Cell wall structure maintenance | [90] | |
| AtFLA1 | Stomata, trichome, leaf vasculature, primary root tip and lateral root | Lateral root development and shoot regeneration | [91] | |
| AtFLA4 (SOS5) | Flower, leaf, stem, root, silique | Root salt stress tolerance; seed mucilage adherence | [92][93][94][95][96] | |
| PpAGP1 | Apical cells | Apical cell expansion | [97] | |
| AtAGP30 | Root | Root regeneration and seed germination | [60] | |
| BcrFLA1 | Root | Root hair elongation | [98] | |
| Secondary wall development | AtFLA11, AtFLA12 | Stem and branch | Secondary cell wall synthesis/patterning | [99] |
| AtXYP1, AtXYP2 | Cell walls of differentiating tracheary elements | Vascular tissue development and patterning | [100] | |
| GhAGP4 | Cotton fiber | Cotton fiber initiation and elongation | [81] | |
| GhFLA1 | Cotton fiber | Fiber initiation and elongation | [14] | |
| PtFLA6 | Stem xylem fiber | Secondary cell wall synthesis/patterning | [101] | |
| Defense | SlattAGP | Site of parasite attack | Promotes parasite adherence | [102] |
| Plant-microbe interaction | AtAGP17 | Root | Agrobacterium tumefaciens root transformation | [103] |
- ↑ Gh: w:Gossypium hirsutum, Dc: w:Daucus carota, At: w:Arabidopsis thaliana, Nt: w:Nicotiana tabacum, Bg: w:Bactris gasipaes, Bn: w:Brassica napus, Ma: w:Musa spp. AAA, Ps: w:Pelargonium sidoides, Fs: w:Fucus serratus, Vc: w:Volvox carteri, Bcm: w:Brassica campestris, Np: w:Nicotiana plumbaginifolia, Na: w:Nicotiana alata, Os: w:Oryza sativa, Pp: w:Physcomitrella patens, Bcr: w:Brassica carinata, Pt: w:Populus trichocarpa, Sl: w:Solanum lycopersicum.
- ↑ PLA: phytocyanin-like AGP. ALGAL-CAM: algal cell adhesion molecule. ISG: inversion-specific glycoprotein. FLA: fasciclin like AGP. ENODL: earlt nodulation like. MF: male fertility. TTS: transmitting tissue specific. MTR: microspore and tapetum regulator. SOS: salt overly sensitive. XYP: xylogen protein. attAGP: attachment AGP.
Additional information
Acknowledgements
The authors would like to acknowledge the support of the La Trobe Institute for Agriculture and Food and a La Trobe Research Focus Area grant 2000004372.
Competing interests
The authors have no competing interests to declare.
Ethics statement
No animal or human research was performed.
References
- ↑ 1.0 1.1 1.2 Showalter, Allan M.; Keppler, Brian; Lichtenberg, Jens; Gu, Dazhang; Welch, Lonnie R. (2010-04-15). "A Bioinformatics Approach to the Identification, Classification, and Analysis of Hydroxyproline-Rich Glycoproteins". Plant Physiology 153 (2): 485–513. doi:10.1104/pp.110.156554. ISSN 0032-0889. http://dx.doi.org/10.1104/pp.110.156554.
- ↑ 2.0 2.1 2.2 2.3 2.4 2.5 Johnson, Kim L.; Cassin, Andrew M.; Lonsdale, Andrew; Bacic, Antony; Doblin, Monika S.; Schultz, Carolyn J. (2017-04-26). "Pipeline to Identify Hydroxyproline-Rich Glycoproteins". Plant Physiology 174 (2): 886–903. doi:10.1104/pp.17.00294. ISSN 0032-0889. http://dx.doi.org/10.1104/pp.17.00294.
- ↑ 3.0 3.1 3.2 3.3 3.4 3.5 Johnson, Kim L.; Cassin, Andrew M.; Lonsdale, Andrew; Wong, Gane Ka-Shu; Soltis, Douglas E.; Miles, Nicholas W.; Melkonian, Michael; Melkonian, Barbara et al. (2017-04-26). "Insights into the Evolution of Hydroxyproline-Rich Glycoproteins from 1000 Plant Transcriptomes". Plant Physiology 174 (2): 904–921. doi:10.1104/pp.17.00295. ISSN 0032-0889. http://dx.doi.org/10.1104/pp.17.00295.
- ↑ 4.0 4.1 4.2 4.3 Schultz, Carolyn J.; Rumsewicz, Michael P.; Johnson, Kim L.; Jones, Brian J.; Gaspar, Yolanda M.; Bacic, Antony (2002-08-01). "Using Genomic Resources to Guide Research Directions. The Arabinogalactan Protein Gene Family as a Test Case". Plant Physiology 129 (4): 1448–1463. doi:10.1104/pp.003459. ISSN 0032-0889. http://dx.doi.org/10.1104/pp.003459.
- ↑ 5.0 5.1 Ma, Yingxuan; Zeng, Wei; Bacic, Antony; Johnson, Kim (2018). "AGPs Through Time and Space". Annual Plant Reviews Online 3 (1): 767–804. doi:10.1002/9781119312994.apr0608. ISSN 2639-3832. https://onlinelibrary.wiley.com/doi/abs/10.1002/9781119312994.apr0608.
- ↑ Shafee, Thomas; Bacic, Antony; Johnson, Kim (2020-08-01). Wilke, Claus. ed. "Evolution of Sequence-Diverse Disordered Regions in a Protein Family: Order within the Chaos". Molecular Biology and Evolution 37 (8): 2155–2172. doi:10.1093/molbev/msaa096. ISSN 0737-4038. https://academic.oup.com/mbe/article/37/8/2155/5828228.
- ↑ Kieliszewski, Marcia J.; Lamport, Derek T.A. (1994-02). "Extensin: repetitive motifs, functional sites, post-translational codes, and phylogeny". The Plant Journal 5 (2): 157–172. doi:10.1046/j.1365-313x.1994.05020157.x. ISSN 0960-7412. http://dx.doi.org/10.1046/j.1365-313x.1994.05020157.x.
- ↑ Tan, Li; Leykam, Joseph F.; Kieliszewski, Marcia J. (2003-06-12). "Glycosylation Motifs That Direct Arabinogalactan Addition to Arabinogalactan-Proteins". Plant Physiology 132 (3): 1362–1369. doi:10.1104/pp.103.021766. ISSN 0032-0889. http://dx.doi.org/10.1104/pp.103.021766.
- ↑ Schultz, Carolyn; Gilson, Paul; Oxley, David; Youl, Joelian; Bacic, Antony (1998-11). "GPI-anchors on arabinogalactan-proteins: implications for signalling in plants". Trends in Plant Science 3 (11): 426–431. doi:10.1016/s1360-1385(98)01328-4. ISSN 1360-1385. http://dx.doi.org/10.1016/s1360-1385(98)01328-4.
- ↑ Koski, M. Kristian; Hieta, Reija; Böllner, Claudia; Kivirikko, Kari I.; Myllyharju, Johanna; Wierenga, Rik K. (2007-12-21). "The active site of an algal prolyl 4-hydroxylase has a large structural plasticity". The Journal of Biological Chemistry 282 (51): 37112–37123. doi:10.1074/jbc.M706554200. ISSN 0021-9258. PMID 17940281. https://www.ncbi.nlm.nih.gov/pubmed/17940281.
- ↑ 11.0 11.1 11.2 11.3 11.4 Ellis, Miriam; Egelund, Jack; Schultz, Carolyn J.; Bacic, Antony (2010-04-13). "Arabinogalactan-Proteins: Key Regulators at the Cell Surface?". Plant Physiology 153 (2): 403–419. doi:10.1104/pp.110.156000. ISSN 0032-0889. http://dx.doi.org/10.1104/pp.110.156000.
- ↑ Showalter, A.M. (2001-09). "Arabinogalactan-proteins: structure, expression and function". Cellular and Molecular Life Sciences 58 (10): 1399–1417. doi:10.1007/pl00000784. ISSN 1420-682X. http://dx.doi.org/10.1007/pl00000784.
- ↑ Gaspar, Yolanda Maria; Nam, Jaesung; Schultz, Carolyn Jane; Lee, Lan-Ying; Gilson, Paul R.; Gelvin, Stanton B.; Bacic, Antony (2004-07-30). "Characterization of the Arabidopsis Lysine-Rich Arabinogalactan-Protein AtAGP17 Mutant (rat1) That Results in a Decreased Efficiency of Agrobacterium Transformation". Plant Physiology 135 (4): 2162–2171. doi:10.1104/pp.104.045542. ISSN 0032-0889. http://dx.doi.org/10.1104/pp.104.045542.
- ↑ 14.0 14.1 Huang, Geng-Qing; Gong, Si-Ying; Xu, Wen-Liang; Li, Wen; Li, Peng; Zhang, Chao-Jun; Li, Deng-Di; Zheng, Yong et al. (2013-01-24). "A Fasciclin-Like Arabinogalactan Protein, GhFLA1, Is Involved in Fiber Initiation and Elongation of Cotton". Plant Physiology 161 (3): 1278–1290. doi:10.1104/pp.112.203760. ISSN 0032-0889. http://dx.doi.org/10.1104/pp.112.203760.
- ↑ 15.0 15.1 Ma, Yuling; Yan, Chenchao; Li, Huimin; Wu, Wentao; Liu, Yaxue; Wang, Yuqian; Chen, Qin; Ma, Haoli (2017-01-26). "Bioinformatics Prediction and Evolution Analysis of Arabinogalactan Proteins in the Plant Kingdom". Frontiers in Plant Science 8. doi:10.3389/fpls.2017.00066. ISSN 1664-462X. PMID 28184232. PMC PMC5266747. http://journal.frontiersin.org/article/10.3389/fpls.2017.00066/full.
- ↑ Dragićević, Milan B; Paunović, Danijela M; Bogdanović, Milica D; .Todorović, Sladjana I; Simonović, Ana D (2020-01-01). "ragp: Pipeline for mining of plant hydroxyproline-rich glycoproteins with implementation in R". Glycobiology 30 (1): 19–35. doi:10.1093/glycob/cwz072. ISSN 1460-2423. https://academic.oup.com/glycob/article/30/1/19/5567434.
- ↑ Pfeifer, Lukas; Shafee, Thomas; Johnson, Kim L.; Bacic, Antony; Classen, Birgit (2020-12). "Arabinogalactan-proteins of Zostera marina L. contain unique glycan structures and provide insight into adaption processes to saline environments". Scientific Reports 10 (1): 8232. doi:10.1038/s41598-020-65135-5. ISSN 2045-2322. PMID 32427862. PMC PMC7237498. http://www.nature.com/articles/s41598-020-65135-5.
- ↑ 18.0 18.1 Baldwin, Timothy C.; Domingo, Concha; Schindler, Thomas; Seetharaman, Gouri; Stacey, Nicola; Roberts, Keith (2001). "DcAGP1, a secreted arabinogalactan protein, is related to a family of basic proline-rich proteins". Plant Molecular Biology 45 (4): 421–435. doi:10.1023/A:1010637426934. http://link.springer.com/10.1023/A:1010637426934.
- ↑ Nguyen-Kim, Huan; San Clemente, Hélène; Laimer, Josef; Lackner, Peter; Gadermaier, Gabriele; Dunand, Christophe; Jamet, Elisabeth (2020-04-03). "The Cell Wall PAC (Proline-Rich, Arabinogalactan Proteins, Conserved Cysteines) Domain-Proteins Are Conserved in the Green Lineage". International Journal of Molecular Sciences 21 (7): 2488. doi:10.3390/ijms21072488. ISSN 1422-0067. PMID 32260156. PMC PMC7177597. https://www.mdpi.com/1422-0067/21/7/2488.
- ↑ 20.0 20.1 Gaspar, Y.; Johnson, K. L.; McKenna, J. A.; Bacic, A.; Schultz, C. J. (2001-09). "The complex structures of arabinogalactan-proteins and the journey towards understanding function". Plant Molecular Biology 47 (1–2): 161–176. doi:10.1023/A:1010683432529. ISSN 0167-4412. PMID 11554470. https://www.ncbi.nlm.nih.gov/pubmed/11554470.
- ↑ 21.0 21.1 Hervé, Cécile; Siméon, Amandine; Jam, Murielle; Cassin, Andrew; Johnson, Kim L.; Salmeán, Armando A.; Willats, William G. T.; Doblin, Monika S. et al. (2015-12-15). "Arabinogalactan proteins have deep roots in eukaryotes: identification of genes and epitopes in brown algae and their role inFucus serratusembryo development". New Phytologist 209 (4): 1428–1441. doi:10.1111/nph.13786. ISSN 0028-646X. http://dx.doi.org/10.1111/nph.13786.
- ↑ Liang, Yan; Basu, Debarati; Pattathil, Sivakumar; Xu, Wen-liang; Venetos, Alexandra; Martin, Stanton L.; Faik, Ahmed; Hahn, Michael G. et al. (2013-10-14). "Biochemical and physiological characterization of fut4 and fut6 mutants defective in arabinogalactan-protein fucosylation in Arabidopsis". Journal of Experimental Botany 64 (18): 5537–5551. doi:10.1093/jxb/ert321. ISSN 1460-2431. http://dx.doi.org/10.1093/jxb/ert321.
- ↑ Oka, Takuji; Saito, Fumie; Shimma, Yoh-ichi; Yoko-o, Takehiko; Nomura, Yoshiyuki; Matsuoka, Ken; Jigami, Yoshifumi (2009-11-18). "Characterization of Endoplasmic Reticulum-Localized UDP-d-Galactose: Hydroxyproline O-Galactosyltransferase Using Synthetic Peptide Substrates in Arabidopsis". Plant Physiology 152 (1): 332–340. doi:10.1104/pp.109.146266. ISSN 0032-0889. http://dx.doi.org/10.1104/pp.109.146266.
- ↑ Kato, Hideaki; Takeuchi, Yoshimi; Tsumuraya, Yoichi; Hashimoto, Yohichi; Nakano, Hirofumi; Kováč, Pavol (2003-02-11). "In vitro biosynthesis of galactans by membrane-bound galactosyltransferase from radish (Raphanus sativus L.) seedlings". Planta 217 (2): 271–282. doi:10.1007/s00425-003-0978-7. ISSN 0032-0935. http://dx.doi.org/10.1007/s00425-003-0978-7.
- ↑ Showalter, Allan M.; Basu, Debarati (2016-06-15). "Extensin and Arabinogalactan-Protein Biosynthesis: Glycosyltransferases, Research Challenges, and Biosensors". Frontiers in Plant Science 7. doi:10.3389/fpls.2016.00814. ISSN 1664-462X. http://dx.doi.org/10.3389/fpls.2016.00814.
- ↑ Egelund, Jack; Obel, Nicolai; Ulvskov, Peter; Geshi, Naomi; Pauly, Markus; Bacic, Antony; Petersen, Bent Larsen (2007-03-31). "Molecular characterization of two Arabidopsis thaliana glycosyltransferase mutants, rra1 and rra2, which have a reduced residual arabinose content in a polymer tightly associated with the cellulosic wall residue". Plant Molecular Biology 64 (4): 439–451. doi:10.1007/s11103-007-9162-y. ISSN 0167-4412. http://dx.doi.org/10.1007/s11103-007-9162-y.
- ↑ 27.0 27.1 Qu, Yongmei; Egelund, Jack; Gilson, Paul R.; Houghton, Fiona; Gleeson, Paul A.; Schultz, Carolyn J.; Bacic, Antony (2008-06-12). "Identification of a novel group of putative Arabidopsis thaliana β-(1,3)-galactosyltransferases". Plant Molecular Biology 68 (1–2): 43–59. doi:10.1007/s11103-008-9351-3. ISSN 0167-4412. http://dx.doi.org/10.1007/s11103-008-9351-3.
- ↑ Basu, Debarati; Tian, Lu; Wang, Wuda; Bobbs, Shauni; Herock, Hayley; Travers, Andrew; Showalter, Allan M. (2015-12). "A small multigene hydroxyproline-O-galactosyltransferase family functions in arabinogalactan-protein glycosylation, growth and development in Arabidopsis". BMC Plant Biology 15 (1). doi:10.1186/s12870-015-0670-7. ISSN 1471-2229. http://dx.doi.org/10.1186/s12870-015-0670-7.
- ↑ Ogawa-Ohnishi, Mari; Matsubayashi, Yoshikatsu (2015-02-24). "Identification of three potent hydroxyprolineO-galactosyltransferases in Arabidopsis". The Plant Journal 81 (5): 736–746. doi:10.1111/tpj.12764. ISSN 0960-7412. http://dx.doi.org/10.1111/tpj.12764.
- ↑ 30.0 30.1 Knoch, Eva; Dilokpimol, Adiphol; Tryfona, Theodora; Poulsen, Christian P.; Xiong, Guangyan; Harholt, Jesper; Petersen, Bent L.; Ulvskov, Peter et al. (2013-11-29). "A β-glucuronosyltransferase fromArabidopsis thalianainvolved in biosynthesis of type II arabinogalactan has a role in cell elongation during seedling growth". The Plant Journal 76 (6): 1016–1029. doi:10.1111/tpj.12353. ISSN 0960-7412. http://dx.doi.org/10.1111/tpj.12353.
- ↑ Dilokpimol, Adiphol; Geshi, Naomi (2014-04-16). "Arabidopsis thalianaglucuronosyltransferase in family GT14". Plant Signaling & Behavior 9 (6): e28891. doi:10.4161/psb.28891. ISSN 1559-2324. http://dx.doi.org/10.4161/psb.28891.
- ↑ Wu, Yingying; Williams, Matthew; Bernard, Sophie; Driouich, Azeddine; Showalter, Allan M.; Faik, Ahmed (2010-04-30). "Functional identification of two nonredundant Arabidopsis alpha(1,2)fucosyltransferases specific to arabinogalactan proteins". The Journal of Biological Chemistry 285 (18): 13638–13645. doi:10.1074/jbc.M110.102715. ISSN 1083-351X. PMID 20194500. PMC 2859526. https://www.ncbi.nlm.nih.gov/pubmed/20194500.
- ↑ Tryfona, Theodora; Theys, Tina E.; Wagner, Tanya; Stott, Katherine; Keegstra, Kenneth; Dupree, Paul (2014-03-25). "Characterisation of FUT4 and FUT6 α-(1→2)-Fucosyltransferases Reveals that Absence of Root Arabinogalactan Fucosylation Increases Arabidopsis Root Growth Salt Sensitivity". PLoS ONE 9 (3): e93291. doi:10.1371/journal.pone.0093291. ISSN 1932-6203. http://dx.doi.org/10.1371/journal.pone.0093291.
- ↑ Dilokpimol, Adiphol; Poulsen, Christian; Vereb, György; Kaneko, Satoshi; Schulz, Alexander; Geshi, Naomi (2014). "Galactosyltransferases from Arabidopsis thaliana in the biosynthesis of type II arabinogalactan: molecular interaction enhances enzyme activity". BMC Plant Biology 14 (1): 90. doi:10.1186/1471-2229-14-90. ISSN 1471-2229. http://dx.doi.org/10.1186/1471-2229-14-90.
- ↑ 35.0 35.1 35.2 35.3 Oxley, D.; Bacic, A. (1999-12-07). "Structure of the glycosylphosphatidylinositol anchor of an arabinogalactan protein from Pyrus communis suspension-cultured cells". Proceedings of the National Academy of Sciences of the United States of America 96 (25): 14246–14251. doi:10.1073/pnas.96.25.14246. ISSN 0027-8424. PMID 10588691. PMC PMC24422. https://www.ncbi.nlm.nih.gov/pubmed/10588691.
- ↑ 36.0 36.1 Yeats, Trevor H.; Bacic, Antony; Johnson, Kim L. (2018-08). "Plant glycosylphosphatidylinositol anchored proteins at the plasma membrane-cell wall nexus: Plant GPI-anchored proteins". Journal of Integrative Plant Biology 60 (8): 649–669. doi:10.1111/jipb.12659. ISSN 1744-7909. http://doi.wiley.com/10.1111/jipb.12659.
- ↑ Imhof, Isabella; Flury, Isabelle; Vionnet, Christine; Roubaty, Carole; Egger, Diane; Conzelmann, Andreas (2004-05-07). "Glycosylphosphatidylinositol (GPI) proteins of Saccharomyces cerevisiae contain ethanolamine phosphate groups on the alpha1,4-linked mannose of the GPI anchor". The Journal of Biological Chemistry 279 (19): 19614–19627. doi:10.1074/jbc.M401873200. ISSN 0021-9258. PMID 14985347. https://www.ncbi.nlm.nih.gov/pubmed/14985347.
- ↑ Ferguson, M.; Homans, S.; Dwek, R.; Rademacher, T. (1988-02-12). "Glycosyl-phosphatidylinositol moiety that anchors Trypanosoma brucei variant surface glycoprotein to the membrane". Science 239 (4841): 753–759. doi:10.1126/science.3340856. ISSN 0036-8075. http://dx.doi.org/10.1126/science.3340856.
- ↑ Ferguson, M. A. (1999-09). "The structure, biosynthesis and functions of glycosylphosphatidylinositol anchors, and the contributions of trypanosome research". Journal of Cell Science 112 (17): 2799–2809. doi:10.1242/jcs.112.17.2799. ISSN 0021-9533. PMID 10444375. https://www.ncbi.nlm.nih.gov/pubmed/10444375.
- ↑ Strasser, Richard (2016-02-23). "Plant protein glycosylation". Glycobiology 26 (9): 926–939. doi:10.1093/glycob/cww023. ISSN 0959-6658. http://dx.doi.org/10.1093/glycob/cww023.
- ↑ Muniz, M.; Zurzolo, C. (2014-06-06). "Sorting of GPI-anchored proteins from yeast to mammals - common pathways at different sites?". Journal of Cell Science 127 (13): 2793–2801. doi:10.1242/jcs.148056. ISSN 0021-9533. http://dx.doi.org/10.1242/jcs.148056.
- ↑ Saha, Dipjyoti; Bhattacharya, Suvendu (2010-12). "Hydrocolloids as thickening and gelling agents in food: a critical review". Journal of Food Science and Technology 47 (6): 587–597. doi:10.1007/s13197-010-0162-6. ISSN 0022-1155. PMID 23572691. PMC PMC3551143. http://link.springer.com/10.1007/s13197-010-0162-6.
- ↑ Barclay, Thomas G.; Day, Candace Minhthu; Petrovsky, Nikolai; Garg, Sanjay (2019-10). "Review of polysaccharide particle-based functional drug delivery". Carbohydrate Polymers 221: 94–112. doi:10.1016/j.carbpol.2019.05.067. PMID 31227171. PMC PMC6626612. https://linkinghub.elsevier.com/retrieve/pii/S0144861719305831.
- ↑ Aizat, Wan M.; Preuss, James M.; Johnson, Alexander A.T.; Tester, Mark A.; Schultz, Carolyn J. (2011-11). "Investigation of a His-rich arabinogalactan-protein for micronutrient biofortification of cereal grain". Physiologia Plantarum 143 (3): 271–286. doi:10.1111/j.1399-3054.2011.01499.x. http://doi.wiley.com/10.1111/j.1399-3054.2011.01499.x.
- ↑ Fujita, Kiyotaka; Sasaki, Yuki; Kitahara, Kanefumi (2019-09). "Degradation of plant arabinogalactan proteins by intestinal bacteria: characteristics and functions of the enzymes involved". Applied Microbiology and Biotechnology 103 (18): 7451–7457. doi:10.1007/s00253-019-10049-0. ISSN 0175-7598. http://link.springer.com/10.1007/s00253-019-10049-0.
- ↑ Singha, Prajjal K.; Roy, Somenath; Dey, Satyahari (2007-04). "Protective activity of andrographolide and arabinogalactan proteins from Andrographis paniculata Nees. against ethanol-induced toxicity in mice". Journal of Ethnopharmacology 111 (1): 13–21. doi:10.1016/j.jep.2006.10.026. https://linkinghub.elsevier.com/retrieve/pii/S037887410600554X.
- ↑ Fincher, G B; Stone, B A; Clarke, A E (1983-06-01). "Arabinogalactan-Proteins: Structure, Biosynthesis, and Function". Annual Review of Plant Physiology 34 (1): 47–70. doi:10.1146/annurev.pp.34.060183.000403. ISSN 0066-4294. https://www.annualreviews.org/doi/10.1146/annurev.pp.34.060183.000403.
- ↑ Nguema-Ona, Eric; Vicré-Gibouin, Maïté; Cannesan, Marc-Antoine; Driouich, Azeddine (2013-08). "Arabinogalactan proteins in root–microbe interactions". Trends in Plant Science 18 (8): 440–449. doi:10.1016/j.tplants.2013.03.006. ISSN 1878-4372. https://linkinghub.elsevier.com/retrieve/pii/S1360138513000599.
- ↑ Filmus, Jorge; Capurro, Mariana; Rast, Jonathan (2008). "Glypicans". Genome Biology 9 (5): 224. doi:10.1186/gb-2008-9-5-224. ISSN 1465-6906. http://dx.doi.org/10.1186/gb-2008-9-5-224.
- ↑ Schaefer, Liliana; Schaefer, Roland M. (2009-06-10). "Proteoglycans: from structural compounds to signaling molecules". Cell and Tissue Research 339 (1): 237–246. doi:10.1007/s00441-009-0821-y. ISSN 0302-766X. http://dx.doi.org/10.1007/s00441-009-0821-y.
- ↑ Tan, Li; Showalter, Allan M.; Egelund, Jack; Hernandez-Sanchez, Arianna; Doblin, Monika S.; Bacic, Antony (2012). "Arabinogalactan-proteins and the research challenges for these enigmatic plant cell surface proteoglycans". Frontiers in Plant Science 3. doi:10.3389/fpls.2012.00140. ISSN 1664-462X. http://dx.doi.org/10.3389/fpls.2012.00140.
- ↑ Kitazawa, Kiminari; Tryfona, Theodora; Yoshimi, Yoshihisa; Hayashi, Yoshihiro; Kawauchi, Susumu; Antonov, Liudmil; Tanaka, Hiroshi; Takahashi, Takashi et al. (2013-03). "β-Galactosyl Yariv Reagent Binds to the β-1,3-Galactan of Arabinogalactan Proteins". Plant Physiology 161 (3): 1117–1126. doi:10.1104/pp.112.211722. ISSN 0032-0889. PMID 23296690. PMC PMC3585584. http://www.plantphysiol.org/lookup/doi/10.1104/pp.112.211722.
- ↑ Yariv, J; Rapport, MM; Graf, L (1962-11-01). "The interaction of glycosides and saccharides with antibody to the corresponding phenylazo glycosides". Biochemical Journal 85 (2): 383–388. doi:10.1042/bj0850383. ISSN 0006-2936. http://dx.doi.org/10.1042/bj0850383.
- ↑ 54.0 54.1 Tang, X.-C. (2006-07-07). "The role of arabinogalactan proteins binding to Yariv reagents in the initiation, cell developmental fate, and maintenance of microspore embryogenesis in Brassica napus L. cv. Topas". Journal of Experimental Botany 57 (11): 2639–2650. doi:10.1093/jxb/erl027. ISSN 0022-0957. http://dx.doi.org/10.1093/jxb/erl027.
- ↑ Willats, William G.T.; Knox, J. Paul (1996-06). "A role for arabinogalactan-proteins in plant cell expansion: evidence from studies on the interaction of beta-glucosyl Yariv reagent with seedlings of Arabidopsis thaliana". The Plant Journal 9 (6): 919–925. doi:10.1046/j.1365-313x.1996.9060919.x. ISSN 0960-7412. http://dx.doi.org/10.1046/j.1365-313x.1996.9060919.x.
- ↑ Chapman, Audrey; Blervacq, Anne-Sophie; Vasseur, Jacques; Hilbert, Jean-Louis (2000-08-10). "Arabinogalactan-proteins in Cichorium somatic embryogenesis: effect of β-glucosyl Yariv reagent and epitope localisation during embryo development". Planta 211 (3): 305–314. doi:10.1007/s004250000299. ISSN 0032-0935. http://dx.doi.org/10.1007/s004250000299.
- ↑ Zagorchev, L; Stoineva, R; Odjakova, M (2013). "Changes in arabinogalactan proteins during somatic embryogenesis In suspension In vitro Cultures of Dactylis glomerata L.". Bulgarian Journal of Agricultural Science 17 (2): 35–38. ISSN 1310-0351. http://www.agrojournal.org/19/02-08s.pdf.
- ↑ Ruprecht, Colin; Bartetzko, Max P.; Senf, Deborah; Dallabernadina, Pietro; Boos, Irene; Andersen, Mathias C.F.; Kotake, Toshihisa; Knox, J. Paul et al. (2017-11). "A Synthetic Glycan Microarray Enables Epitope Mapping of Plant Cell Wall Glycan-Directed Antibodies". Plant Physiology 175 (3): 1094–1104. doi:10.1104/pp.17.00737. ISSN 0032-0889. PMID 28924016. PMC PMC5664464. http://www.plantphysiol.org/lookup/doi/10.1104/pp.17.00737.
- ↑ Seifert, Georg J.; Roberts, Keith (2007-06). "The Biology of Arabinogalactan Proteins". Annual Review of Plant Biology 58 (1): 137–161. doi:10.1146/annurev.arplant.58.032806.103801. ISSN 1543-5008. http://dx.doi.org/10.1146/annurev.arplant.58.032806.103801.
- ↑ 60.0 60.1 van Hengel, Arjon J.; Roberts, Keith (2003-10). "AtAGP30, an arabinogalactan-protein in the cell walls of the primary root, plays a role in root regeneration and seed germination". The Plant Journal 36 (2): 256–270. doi:10.1046/j.1365-313x.2003.01874.x. ISSN 0960-7412. http://dx.doi.org/10.1046/j.1365-313x.2003.01874.x.
- ↑ Coimbra, Sílvia; Costa, Mário; Mendes, Marta Adelina; Pereira, Ana Marta; Pinto, João; Pereira, Luís Gustavo (2010-02-17). "Early germination of Arabidopsis pollen in a double null mutant for the arabinogalactan protein genes AGP6 and AGP11". Sexual Plant Reproduction 23 (3): 199–205. doi:10.1007/s00497-010-0136-x. ISSN 0934-0882. http://dx.doi.org/10.1007/s00497-010-0136-x.
- ↑ Suzuki, Toshiya; Narciso, Joan Oñate; Zeng, Wei; van de Meene, Allison; Yasutomi, Masayuki; Takemura, Shunsuke; Lampugnani, Edwin R.; Doblin, Monika S. et al. (2016-11-09). "KNS4/UPEX1: A Type II Arabinogalactan β-(1,3)-Galactosyltransferase Required for Pollen Exine Development". Plant Physiology 173 (1): 183–205. doi:10.1104/pp.16.01385. ISSN 0032-0889. http://dx.doi.org/10.1104/pp.16.01385.
- ↑ Lamport, Derek T. A.; Várnai, Péter (2013-01). "Periplasmic arabinogalactan glycoproteins act as a calcium capacitor that regulates plant growth and development". New Phytologist 197 (1): 58–64. doi:10.1111/nph.12005. ISSN 0028-646X. http://doi.wiley.com/10.1111/nph.12005.
- ↑ Lamport, Derek T. A.; Tan, Li; Held, Michael; Kieliszewski, Marcia J. (2020-02-09). "Phyllotaxis Turns Over a New Leaf—A New Hypothesis". International Journal of Molecular Sciences 21 (3): 1145. doi:10.3390/ijms21031145. ISSN 1422-0067. PMID 32050457. PMC PMC7037126. https://www.mdpi.com/1422-0067/21/3/1145.
- ↑ Lopez-Hernandez, Federico; Tryfona, Theodora; Rizza, Annalisa; Yu, Xiaolan L.; Harris, Matthew O.B.; Webb, Alex A.R.; Kotake, Toshihisa; Dupree, Paul (2020-10). "Calcium Binding by Arabinogalactan Polysaccharides Is Important for Normal Plant Development". The Plant Cell 32 (10): 3346–3369. doi:10.1105/tpc.20.00027. ISSN 1040-4651. http://www.plantcell.org/lookup/doi/10.1105/tpc.20.00027.
- ↑ 66.0 66.1 Huber, O.; Sumper, M. (1994-09-15). "Algal-CAMs: isoforms of a cell adhesion molecule in embryos of the alga Volvox with homology to Drosophila fasciclin I". The EMBO Journal 13 (18): 4212–4222. doi:10.1002/j.1460-2075.1994.tb06741.x. ISSN 0261-4189. PMID 7925267. PMC PMC395348. https://www.ncbi.nlm.nih.gov/pubmed/7925267.
- ↑ Seifert, Georg J. (2018-05-31). "Fascinating Fasciclins: A Surprisingly Widespread Family of Proteins that Mediate Interactions between the Cell Exterior and the Cell Surface". International Journal of Molecular Sciences 19 (6). doi:10.3390/ijms19061628. ISSN 1422-0067. PMID 29857505. PMC 6032426. https://www.ncbi.nlm.nih.gov/pubmed/29857505.
- ↑ Poon, Simon; Heath, Robyn Louise; Clarke, Adrienne Elizabeth (2012-08-02). "A Chimeric Arabinogalactan Protein Promotes Somatic Embryogenesis in Cotton Cell Culture". Plant Physiology 160 (2): 684–695. doi:10.1104/pp.112.203075. ISSN 0032-0889. http://dx.doi.org/10.1104/pp.112.203075.
- ↑ Toonen, Marcel A. J.; Schmidt, Ed D. L.; van Kammen, Ab; de Vries, Sacco C. (1997-09-26). "Promotive and inhibitory effects of diverse arabinogalactan proteins on Daucus carota L. somatic embryogenesis". Planta 203 (2): 188–195. doi:10.1007/s004250050181. ISSN 0032-0935. http://dx.doi.org/10.1007/s004250050181.
- ↑ Hu, Ying; Qin, Yuan; Zhao, Jie (2006-10-06). "Localization of an arabinogalactan protein epitope and the effects of Yariv phenylglycoside during zygotic embryo development of Arabidopsis thaliana". Protoplasma 229 (1): 21–31. doi:10.1007/s00709-006-0185-z. ISSN 0033-183X. http://dx.doi.org/10.1007/s00709-006-0185-z.
- ↑ Qin, Y. (2006-01-31). "Localization of arabinogalactan proteins in egg cells, zygotes, and two-celled proembryos and effects of -D-glucosyl Yariv reagent on egg cell fertilization and zygote division in Nicotiana tabacum L.". Journal of Experimental Botany 57 (9): 2061–2074. doi:10.1093/jxb/erj159. ISSN 0022-0957. http://dx.doi.org/10.1093/jxb/erj159.
- ↑ Steinmacher, Douglas A.; Saare-Surminski, Katja; Lieberei, Reinhard (2012-06-19). "Arabinogalactan proteins and the extracellular matrix surface network during peach palm somatic embryogenesis". Physiologia Plantarum 146 (3): 336–349. doi:10.1111/j.1399-3054.2012.01642.x. ISSN 0031-9317. http://dx.doi.org/10.1111/j.1399-3054.2012.01642.x.
- ↑ Pan, Xiao; Yang, Xiao; Lin, Guimei; Zou, Ru; Chen, Houbin; Šamaj, Jozef; Xu, Chunxiang (2011-05-24). "Ultrastructural changes and the distribution of arabinogalactan proteins during somatic embryogenesis of banana (Musa spp. AAA cv. ‘Yueyoukang 1’)". Physiologia Plantarum 142 (4): 372–389. doi:10.1111/j.1399-3054.2011.01478.x. ISSN 0031-9317. http://dx.doi.org/10.1111/j.1399-3054.2011.01478.x.
- ↑ Duchow, Stefanie; Dahlke, Renate I.; Geske, Thomas; Blaschek, Wolfgang; Classen, Birgit (2016-11). "Arabinogalactan-proteins stimulate somatic embryogenesis and plant propagation of Pelargonium sidoides". Carbohydrate Polymers 152: 149–155. doi:10.1016/j.carbpol.2016.07.015. ISSN 0144-8617. http://dx.doi.org/10.1016/j.carbpol.2016.07.015.
- ↑ Hallmann, A.; Kirk, D. L. (2000-12). "The developmentally regulated ECM glycoprotein ISG plays an essential role in organizing the ECM and orienting the cells of Volvox". Journal of Cell Science 113 (24): 4605–4617. doi:10.1242/jcs.113.24.4605. ISSN 0021-9533. PMID 11082052. https://www.ncbi.nlm.nih.gov/pubmed/11082052.
- ↑ Pereira, Ana Marta; Lopes, Ana Lúcia; Coimbra, Sílvia (2016-07-14). "JAGGER, an AGP essential for persistent synergid degeneration and polytubey block in Arabidopsis". Plant Signaling & Behavior 11 (8): e1209616. doi:10.1080/15592324.2016.1209616. ISSN 1559-2324. http://dx.doi.org/10.1080/15592324.2016.1209616.
- ↑ Levitin, Bella; Richter, Dganit; Markovich, Inbal; Zik, Moriyah (2008-11). "Arabinogalactan proteins 6 and 11 are required for stamen and pollen function in Arabidopsis". The Plant Journal 56 (3): 351–363. doi:10.1111/j.1365-313x.2008.03607.x. ISSN 0960-7412. http://dx.doi.org/10.1111/j.1365-313x.2008.03607.x.
- ↑ Coimbra, S.; Costa, M.; Jones, B.; Mendes, M. A.; Pereira, L. G. (2009-05-11). "Pollen grain development is compromised in Arabidopsis agp6 agp11 null mutants". Journal of Experimental Botany 60 (11): 3133–3142. doi:10.1093/jxb/erp148. ISSN 0022-0957. http://dx.doi.org/10.1093/jxb/erp148.
- ↑ Acosta-García, Gerardo; Vielle-Calzada, Jean-Philippe (2004-09-17). "A Classical Arabinogalactan Protein Is Essential for the Initiation of Female Gametogenesis in Arabidopsis". The Plant Cell 16 (10): 2614–2628. doi:10.1105/tpc.104.024588. ISSN 1040-4651. http://dx.doi.org/10.1105/tpc.104.024588.
- ↑ Demesa-Arévalo, Edgar; Vielle-Calzada, Jean-Philippe (2013-04). "The Classical Arabinogalactan Protein AGP18 Mediates Megaspore Selection in Arabidopsis". The Plant Cell 25 (4): 1274–1287. doi:10.1105/tpc.112.106237. ISSN 1040-4651. http://dx.doi.org/10.1105/tpc.112.106237.
- ↑ 81.0 81.1 Li, Yunjing; Liu, Diqiu; Tu, Lili; Zhang, Xianlong; Wang, Li; Zhu, Longfu; Tan, Jiafu; Deng, Fenglin (2009-12-30). "Suppression of GhAGP4 gene expression repressed the initiation and elongation of cotton fiber". Plant Cell Reports 29 (2): 193–202. doi:10.1007/s00299-009-0812-1. ISSN 0721-7714. http://dx.doi.org/10.1007/s00299-009-0812-1.
- ↑ Mashiguchi, Kiyoshi; Asami, Tadao; Suzuki, Yoshihito (2009-11-23). "Genome-Wide Identification, Structure and Expression Studies, and Mutant Collection of 22 Early Nodulin-Like Protein Genes in Arabidopsis". Bioscience, Biotechnology, and Biochemistry 73 (11): 2452–2459. doi:10.1271/bbb.90407. ISSN 0916-8451. http://dx.doi.org/10.1271/bbb.90407.
- ↑ Hou, Yingnan; Guo, Xinyang; Cyprys, Philipp; Zhang, Ying; Bleckmann, Andrea; Cai, Le; Huang, Qingpei; Luo, Yu et al. (2016-09). "Maternal ENODLs Are Required for Pollen Tube Reception in Arabidopsis". Current Biology 26 (17): 2343–2350. doi:10.1016/j.cub.2016.06.053. ISSN 0960-9822. http://dx.doi.org/10.1016/j.cub.2016.06.053.
- ↑ Lin, Sue; Dong, Heng; Zhang, Fang; Qiu, Lin; Wang, Fangzhan; Cao, Jiashu; Huang, Li (2014-01-31). "BcMF8, a putative arabinogalactan protein-encoding gene, contributes to pollen wall development, aperture formation and pollen tube growth in Brassica campestris". Annals of Botany 113 (5): 777–788. doi:10.1093/aob/mct315. ISSN 1095-8290. http://dx.doi.org/10.1093/aob/mct315.
- ↑ Lin, Sue; Yue, Xiaoyan; Miao, Yingjing; Yu, Youjian; Dong, Heng; Huang, Li; Cao, Jiashu (2018-03-09). "The distinct functions of two classical arabinogalactan proteins BcMF8 and BcMF18 during pollen wall development in Brassica campestris". The Plant Journal 94 (1): 60–76. doi:10.1111/tpj.13842. ISSN 0960-7412. http://dx.doi.org/10.1111/tpj.13842.
- ↑ Cheung, Alice Y; Wang, Hong; Wu, Hen-ming (1995-08). "A floral transmitting tissue-specific glycoprotein attracts pollen tubes and stimulates their growth". Cell 82 (3): 383–393. doi:10.1016/0092-8674(95)90427-1. ISSN 0092-8674. http://dx.doi.org/10.1016/0092-8674(95)90427-1.
- ↑ Nathan Hancock, C.; Kent, Lia; McClure, Bruce A. (2005-08-08). "The stylar 120 kDa glycoprotein is required for S-specific pollen rejection in Nicotiana". The Plant Journal 43 (5): 716–723. doi:10.1111/j.1365-313x.2005.02490.x. ISSN 0960-7412. http://dx.doi.org/10.1111/j.1365-313x.2005.02490.x.
- ↑ Tan, Hexin; Liang, Wanqi; Hu, Jianping; Zhang, Dabing (2012-06). "MTR1 Encodes a Secretory Fasciclin Glycoprotein Required for Male Reproductive Development in Rice". Developmental Cell 22 (6): 1127–1137. doi:10.1016/j.devcel.2012.04.011. ISSN 1534-5807. http://dx.doi.org/10.1016/j.devcel.2012.04.011.
- ↑ Yang, Jie; Sardar, Harjinder S.; McGovern, Kathleen R.; Zhang, Yizhu; Showalter, Allan M. (2007-01-08). "A lysine-rich arabinogalactan protein in Arabidopsis is essential for plant growth and development, including cell division and expansion". The Plant Journal 49 (4): 629–640. doi:10.1111/j.1365-313x.2006.02985.x. ISSN 0960-7412. http://dx.doi.org/10.1111/j.1365-313x.2006.02985.x.
- ↑ Tan, Li; Eberhard, Stefan; Pattathil, Sivakumar; Warder, Clayton; Glushka, John; Yuan, Chunhua; Hao, Zhangying; Zhu, Xiang et al. (2013-01). "An Arabidopsis Cell Wall Proteoglycan Consists of Pectin and Arabinoxylan Covalently Linked to an Arabinogalactan Protein". The Plant Cell 25 (1): 270–287. doi:10.1105/tpc.112.107334. ISSN 1040-4651. http://dx.doi.org/10.1105/tpc.112.107334.
- ↑ Johnson, Kim L.; Kibble, Natalie A. J.; Bacic, Antony; Schultz, Carolyn J. (2011-09-22). "A Fasciclin-Like Arabinogalactan-Protein (FLA) Mutant of Arabidopsis thaliana, fla1, Shows Defects in Shoot Regeneration". PLoS ONE 6 (9): e25154. doi:10.1371/journal.pone.0025154. ISSN 1932-6203. http://dx.doi.org/10.1371/journal.pone.0025154.
- ↑ Shi, Huazhong; Kim, YongSig; Guo, Yan; Stevenson, Becky; Zhu, Jian-Kang (2002-12-13). "The Arabidopsis SOS5 Locus Encodes a Putative Cell Surface Adhesion Protein and Is Required for Normal Cell Expansion". The Plant Cell 15 (1): 19–32. doi:10.1105/tpc.007872. ISSN 1040-4651. http://dx.doi.org/10.1105/tpc.007872.
- ↑ Harpaz-Saad, Smadar; McFarlane, Heather E.; Xu, Shouling; Divi, Uday K.; Forward, Bronwen; Western, Tamara L.; Kieber, Joseph J. (2011-10-10). "Cellulose synthesis via the FEI2 RLK/SOS5 pathway and CELLULOSE SYNTHASE 5 is required for the structure of seed coat mucilage in Arabidopsis". The Plant Journal 68 (6): 941–953. doi:10.1111/j.1365-313x.2011.04760.x. ISSN 0960-7412. http://dx.doi.org/10.1111/j.1365-313x.2011.04760.x.
- ↑ Griffiths, Jonathan S.; Tsai, Allen Yi-Lun; Xue, Hui; Voiniciuc, Cătălin; Šola, Krešimir; Seifert, Georg J.; Mansfield, Shawn D.; Haughn, George W. (2014-05-07). "SALT-OVERLY SENSITIVE5 Mediates Arabidopsis Seed Coat Mucilage Adherence and Organization through Pectins". Plant Physiology 165 (3): 991–1004. doi:10.1104/pp.114.239400. ISSN 0032-0889.
- ↑ Griffiths, Jonathan S.; Crepeau, Marie-Jeanne; Ralet, Marie-Christine; Seifert, Georg J.; North, Helen M. (2016-07-29). "Dissecting Seed Mucilage Adherence Mediated by FEI2 and SOS5". Frontiers in Plant Science 7. doi:10.3389/fpls.2016.01073. ISSN 1664-462X. http://dx.doi.org/10.3389/fpls.2016.01073.
- ↑ Xue, Hui; Veit, Christiane; Abas, Lindy; Tryfona, Theodora; Maresch, Daniel; Ricardi, Martiniano M.; Estevez, José Manuel; Strasser, Richard et al. (2017-06-13). "Arabidopsis thaliana FLA4 functions as a glycan-stabilized soluble factor via its carboxy-proximal Fasciclin 1 domain". The Plant Journal 91 (4): 613–630. doi:10.1111/tpj.13591. ISSN 0960-7412.
- ↑ Lee, Kieran J.D.; Sakata, Yoichi; Mau, Shaio-Lim; Pettolino, Filomena; Bacic, Antony; Quatrano, Ralph S.; Knight, Celia D.; Knox, J. Paul (2005-09-30). "Arabinogalactan Proteins Are Required for Apical Cell Extension in the Moss Physcomitrella patens". The Plant Cell 17 (11): 3051–3065. doi:10.1105/tpc.105.034413. ISSN 1040-4651. http://dx.doi.org/10.1105/tpc.105.034413.
- ↑ Kirchner, Thomas W.; Niehaus, Markus; Debener, Thomas; Schenk, Manfred K.; Herde, Marco (2017-09-22). "Efficient generation of mutations mediated by CRISPR/Cas9 in the hairy root transformation system of Brassica carinata". PLOS ONE 12 (9): e0185429. doi:10.1371/journal.pone.0185429. ISSN 1932-6203. http://dx.doi.org/10.1371/journal.pone.0185429.
- ↑ MacMillan, Colleen P.; Mansfield, Shawn D.; Stachurski, Zbigniew H.; Evans, Rob; Southerton, Simon G. (2010-02-24). "Fasciclin-like arabinogalactan proteins: specialization for stem biomechanics and cell wall architecture in Arabidopsis and Eucalyptus". The Plant Journal 62 (4): 689–703. doi:10.1111/j.1365-313x.2010.04181.x. ISSN 0960-7412. http://dx.doi.org/10.1111/j.1365-313x.2010.04181.x.
- ↑ Motose, Hiroyasu; Sugiyama, Munetaka; Fukuda, Hiroo (2004-06). "A proteoglycan mediates inductive interaction during plant vascular development". Nature 429 (6994): 873–878. doi:10.1038/nature02613. ISSN 0028-0836. http://dx.doi.org/10.1038/nature02613.
- ↑ Wang, Haihai; Jiang, Chunmei; Wang, Cuiting; Yang, Yang; Yang, Lei; Gao, Xiaoyan; Zhang, Hongxia (2014-11-26). "Antisense expression of the fasciclin-like arabinogalactan protein FLA6 gene in Populus inhibits expression of its homologous genes and alters stem biomechanics and cell wall composition in transgenic trees". Journal of Experimental Botany 66 (5): 1291–1302. doi:10.1093/jxb/eru479. ISSN 1460-2431. http://dx.doi.org/10.1093/jxb/eru479.
- ↑ Albert, Markus; Belastegui-Macadam, Xana; Kaldenhoff, Ralf (2006-11). "An attack of the plant parasite Cuscuta reflexa induces the expression of attAGP, an attachment protein of the host tomato". The Plant Journal 48 (4): 548–556. doi:10.1111/j.1365-313x.2006.02897.x. ISSN 0960-7412. http://dx.doi.org/10.1111/j.1365-313x.2006.02897.x.
- ↑ Gaspar, Yolanda Maria; Nam, Jaesung; Schultz, Carolyn Jane; Lee, Lan-Ying; Gilson, Paul R.; Gelvin, Stanton B.; Bacic, Antony (2004-08). "Characterization of the Arabidopsis Lysine-Rich Arabinogalactan-Protein AtAGP17 Mutant ( rat1 ) That Results in a Decreased Efficiency of Agrobacterium Transformation". Plant Physiology 135 (4): 2162–2171. doi:10.1104/pp.104.045542. ISSN 0032-0889. PMID 15286287. PMC PMC520787. http://www.plantphysiol.org/lookup/doi/10.1104/pp.104.045542.